Our Focus Now
Currently, our research effort focusses on two areas of interest, run in parallel.
Cytoskeleton structure and function: Katnal2, a multitasking katanin-like protein in vertebrates
A few years ago, our group discovered a novel family of katanin-like proteins (KATNAL2) and provided its functional characterization in mouse cells (Ververis et al., 2016, CMLS). We demonstrated that KATNAL2 proteins are multitasking proteins which are important in microtubule-based processes including cytokinesis, ciliogenesis and centriole dynamics. Additionally, we showed that the activity of KATNAL2 impacts on cell cycle progression through microtubule acetylation and profoundly affects cellular size which is strikingly enlarged when KATNAL2 is silenced.



We subsequently harnessed the power of CRISPR-Cas9, in collaboration with the UC Berkeley, to investigate what happens to vertebrate embryonic development in the absence of katnal2 expression. In the Xenopus tropicalis embryo/tadpole, gene editing of katnal2 had marked effects on neurogenesis, affecting proper blastopore closure, neural tube closure and inhibiting neural crest cell migration. Strikingly, katnal2 knockdown resulted in reduced brain size, particularly in the telencephalic region, accompanied by significant reduction in the number of ventricular zone progenitor cells. Organogenesis was also impacted and, additionally, targeted multiciliated cells showed reduced ciliation, and those fewer cilia appeared shorter and displayed aberrant basal body behavior and apical actin formation (Willsey et al., 2018, Dev Biol).
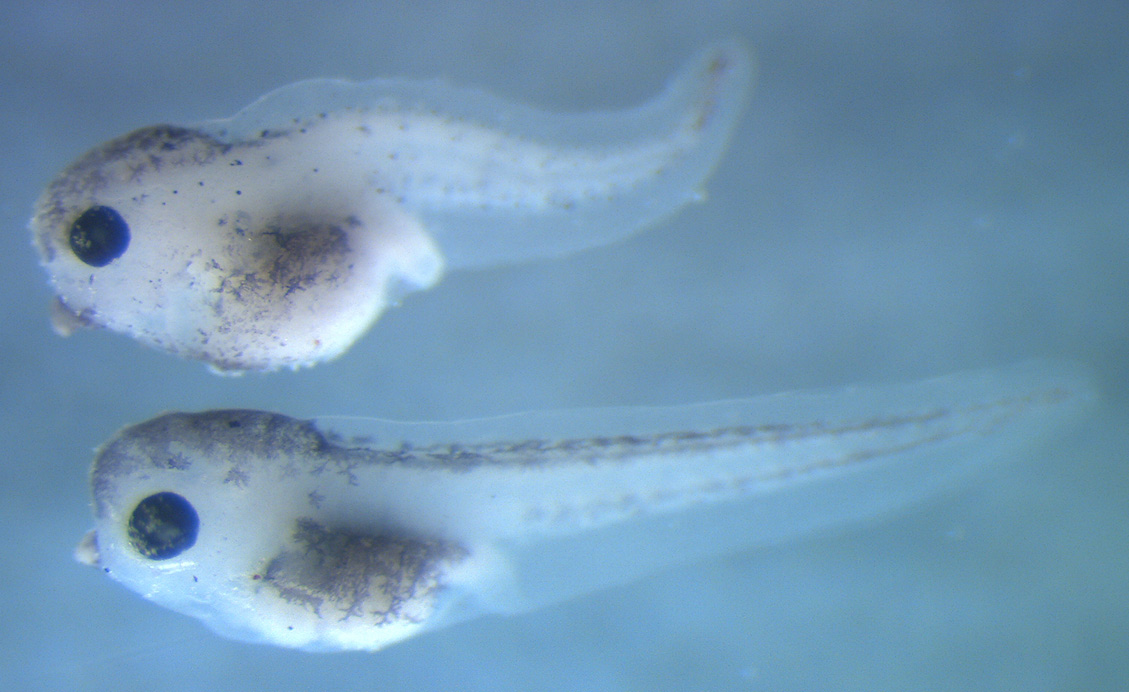


Currently, intense interest in KATNAL2 resulted from several large whole-exome sequencing studies of families that revealed a small number of genes with de novo mutations specifically associated with sporadic Autism Spectrum Disorders and those included the gene for KATNAL2. There is still no understanding of what molecular mechanisms may be involved in the case of KATNAL2 as a factor affecting neurodevelopmental processes pertinent to autism.
We are continuing our work on KATNAL2 and have evidence on yet further critical functions that we are currently characterising and hope to contribute to our further understanding of its impact in cellular physiology.
Endoplasmic Reticulum transmembrane protein function
We have a long-standing interest in ER protein function on the basis of earlier work, in collaboration with Hanry Yu’s group (University of Sungapore), on kinectins in mouse cells (Santama et al., 2004, JCS). Understanding ER morphogenesis, as the ER assembles and disassembles during the cell cycle, is one great challenge. Cell biologists have begun to decipher aspects of how the ER structure is made, they have identified ER proteins that work as wedges, creating membrane curvature, or as scaffolds, bringing membrane sheets in proximity, or as bridges that keep luminal sheets at a constant separation, or creating three-way tubule junctions. In our Christodoulou et al., 2016 JCS study we identified TMEM170A as a novel ER/nuclear envelope protein that functions specifically in shaping ER sheets. We found TMEM170A to physically interact with RTN4, a well-known tubular ER shaping protein, whose overexpression results in ER tubule proliferation. Our experiments showed that the antagonistic activities of TMEM170A and RTN4 are in a functional “tug of war”, where TMEM170A appears to shift the balance towards ER sheet formation while RTN4 promotes tubular ER production.



In our new work by Christodoulou et al., 2020 JCS, we turned our attention on the functional characterization of ER/nuclear envelope transmembrane protein TMEM147. We found that TMEM147 has an intricate relationship with the Lamin B receptor, LBR, which a fascinating protein in its own right, possessing a modular structure and corresponding bifunctionality. LBR, an inner nuclear membrane protein, was initially characterized as a heterochromatin organizer in the nucleus via its N-terminal domain, but more recently also shown to, unexpectedly, exhibit sterol reductase activity at its C-terminus. Silencing of TMEM147 drastically reduces the level LBR at the inner nuclear membrane and results in mistargeting of LBR to the ER. We show that TMEM147 physically interacts with LBR and the C-terminus of LBR is essential for their functional interaction. We discovered that TMEM147 also physically interacts with DHCR7, another key sterol reductase in cholesterol biosynthesis. Similar to LBR, TMEM147 downregulation results in sharp decline of DHCR protein levels and co-ordinate transcriptional decrease of LBR and DHCR7 expression but not with TM7SF2, a sterol reductase exhibiting some functional complementarity with LBR as they both catalyse the same cholesterol biosynthetic step. Consistently, lipidomic analysis upon TMEM147 silencing identified changes in cellular cholesterol levels, cholesteryl ester levels and profile, and in cellular cholesterol uptake, raising the possibility that TMEM147 is an important new regulator of cholesterol homeostasis in cells.


